1
МЫШЦЫ
мускулатура скелетная и внутренних
органов (висцеральная), обеспечивающая у животных и человека выполнение
ряда важнейших физиологич. функций: перемещение тела или отдельных его
частей в пространстве, кровообращение, дыхание, передвижение пищ. кашицы
в пищеварит. органах, поддержание тонуса сосудов, выделение экскрементов
и т. д. Сократит, функция всех типов M. обусловлена превращением в мышечных
волокнах химич. энергии определённых биохимич. процессов в механич. работу.
Однако сокращение скелетных M. и мускулатуры внутренних органов - лишь
частный случай более общей закономерности - механо-химич. активности живых
структур. По-видимому, в основе самых различных проявлений этой активности
- сокращения хвоста сперматозоида, движения ресничек инфузорий, расхождения
хромосом во время митоза, впрыскивания в бактерию фаговой ДНК и
т. д.- лежит один и тот же молекулярный механизм, связанный с возможностью
изменения конформации или взаимного расположения фибриллярных структур
контрактильных белков.
Типы мышц. Морфологи различают 2 осн. типа
M.: поперечнополосатые мышцы и гладкие мышцы. К первым относится
вся скелетная мускулатура позвоночных животных и человека (см. илл. на
вклейке), обеспечивающая возможность выполнения произвольных движений,
M. языка, верх, трети пищевода и некоторые др., M. сердца (миокард), имеющая
свои особенности (состав белков, характер сокращения и др.), а также M.
членистоногих и нек-рых др. бес-лозвонрчных. К гладким M. принадлежит большая
часть мускулатуры беспозвоночных животных и мышечные слои внутр. органов
и стенок кровеносных сосудов позвоночных животных и человека, обеспечивающие
возможность выполнения ряда важнейших физиологич. функций. Нек-рые гистологи,
изучающие M. беспозвоночных, выделяют и 3-й тип M.- с двойной косой исчерченностью
(см. Мышечная ткань).
Структурными элемент а-м и всех типов M.
являются мышечные волокна (рис. 1). Поперечнополосатые мышечные волокна
в скелетных M. образуют пучки, соединённые друг с другом прослойками соединительной
ткани.
Рис. 1. Поперечнополосатые мышечные волокна
человека: а - отрезки двух волокон; б - поперечный разрез
волокна, миофибрил-лы образуют скопления, разграниченные саркоплазмой;
в
- поперечный разрез волокна с равномерным распределением миофибрилл.
Своими концами мышечные волокна сплетаются
с сухожильными волокнами, через посредство к-рых мышечная тяга передаётся
на кости скелета. Волокна поперечнополосатых M. представляют собой гигантские
многоядерные клетки, диаметр к-рых варьирует от 10 до 100мкм, а
длина часто соответствует длине M., достигая, напр., в нек-рых M. человека
12 см. Волокно покрыто эластичной оболочкой - сарколеммой и состоит
из саркоплазмы, структурными элементами к-рой являются такие органоиды,
как митохондрии, рибосомы, трубочки и пузырьки саркоплазматической сети
и т. н. Т-системы (рис. 2), различные включения и т. д.
Рис. 2. Электронная микрофотография саркоплазматической
сети и Т-системы мышечного волокна.
В саркоплазме обычно в форме пучков расположено
множество нитевидных образований толщиной от 0,5 до неск. мкм - миофибрилл,
обладающих, как и всё волокно в целом, поперечной исчерченностью. Каждая
миофибрилла разделена на неск. сот участков длиной 2,5-3 мкм, наз.
саркомерами. Каждый саркомер, в свою очередь, состоит из чередующихся участков
- дисков, обладающих неодинаковой оптической плотностью и придающих миофибриллам
и мышечному волокну в целом характерную поперечную исчерченность, чётко
обнаруживаемую при наблюдении в фазовоконтрастном микроскопе. Более тёмные
диски обладают способностью к двойному лучепреломлению и наз. анизотропными,
или дисками Л. Более светлые диски не обладают этой способностью и наз.
изотропными, или дисками /. Среднюю часть диска А занимает зона
более слабого двойного лучепреломления - зона H. Диск / делится
на 2 равные части тёмной Z-пластинкой, отграничивающей один саркомер от
другого. В каждом сарко-мере имеется два типа нитей (филамен-тов), состоящих
из мышечных белков: толстые миозиновые и тонкие - акти-
Рис. 3. Строение сар-комера поперечно-полосатого
мышечного волокна А - электронная микрофотография (малое увеличение),
на которой чётко видна структура саркоме-ра: Б - схема сар-комера;
В - электронная микрофотография с высокой разрешающей способностью; Г -
поперечное сечение саркомера на различных уровнях, видно положение толстых
и тонких нитей в различных участках покоящегося саркомера (по X. Хаксли).
Скелетные мышцы человека. 1. Вид спереди:
1 - затылочно лобная; 2 - круговая мышца рта; 3 -подбородочная; 4 - грудино-подъязычная;
S - трапециевидная; 6 - трёхглавая мышца плеча; 7 - прямая мышца живота;
8 -наружная косая мышца живота; 9 - подвздошно-поясничная; 10 - мышца,
натягивающая широкую фасцию; 11 - гребешковая; 12 - длинная приводящая;
13 - портняжная; 14 - тонкая; 15 - прямая мышца бедра; 16 - медиальная
широкая мышца бедра; 17 - боковая широкая мышца бедра; 18 - отводящая большой
палец стопы; 19 - длинный разгибатель пальцев (сухожилие); 20 - длинный
разгибатель пальцев; 21 - передняя большеберцовая; 22 - камбаловидная;
23 - икроножная; 24 - короткий разгибатель большого пальца кисти; 25 -
длинный разгибатель большого пальца кисти; 26 - локтевой сгибатель запястья;
27 - короткий лучевой разгибатель запястья; 28 - разгибатель пальцев; 29
- лучевой сгибатель запястья; 30 - длинный лучевой разгибатель запястья;
31 - плече-лучевая; 32 - трёхглавая мышца плеча; 33 - двуглавая мышца плеча;
34 - передняя зубчатая; 35 - большая грудная; 36 - дельтовидная; 37 - средняя
лестничная; 38 - передняя лестничная; 39 и 40 - грудино-ключично-сосцевидная;
41 - опускающая угол рта; 42 - жевательная; 43 - большая скуловая; 44 -
височная. 2. Вид сзади: 1 - затылочно-лобная; 2 - трапециевидная; 3 - дельтовидная;
4 - трёхглавая мышца плеча; 5 - двуглавая мышца плеча; 6 - круглый пронатор;
7 - длинная ладонная; 8 - лучевой сгибатель запястья; 9 - поверхностный
сгибатель пальцев; 10 - плече-лучевая; 11-локтевой сгибатель запястья;
12 - короткая мышца, отводящая большой палец кисти; 13 и 14 - полуперепончатая;
15 - полусухожильная; 16 - тонкая; 17 - двуглавая мышца бедра; 18 - полуперепончатая;
19 - икроножная; 20 - камбаловидная; 21 - большая ягодичная; 22 - натягивающая
широкую фасцию; 23 - средняя ягодичная; 24 - наружная косая живота; 25
- широчайшая мышца спины; 26 - зубчатая передняя; 27 - большая круглая;
28 - подгребешковая; 29 - малая круглая; 30 - плече-лучевая; 31 - грудино-ключично-сосцевидная;
32 - ремённая мышца головы; 33 - жевательная; 34-полуостистая; 35 - височная.
К ст. Мышцы.
К ст. Насекомоядные растения. Ia и 16.
Жирянка
обыкновенная (Pinguicula vulgaris). 2. Росянка капская (Drosera capensis).
3. Росянка круглолистная (Drosera rotundifolia). 4. Росолист (Drosophyllum
lusitanicum). 5. Саррацения (Sarracenia drummondii). 6. Венерина мухоловка
(Dionea muscipula). 7. Непентес (Nepenthes destillatoria). 8. Пузырчатка
обыкновенная (Utricularia vulgaris). 9. Альдрованда (Aldrovanda vesiculosa).
новые (рис. 3). Неск. иную структуру имеют
гладкие мышечные волокна. Они представляют собой веретенообразные одноядерные
клетки, лишённые поперечной исчерченности. Длина их обычно достигает 50-250
мкм
(в
матке - до 500 мкм), ширина - 4-8 мкм', мио-филаменты в них
обычно не объединены в обособленные миофибриллы, а расположены по длине
волокна в виде множества одиночных актиновых нитей (рис. 4). Упорядоченная
система миозиновых нитей в гладкомышечных клетках отсутствует. В гладкой
мускулатуре моллюсков наиболее важную роль в осуществлении запирательной
функции играют, по-видимому, парамиозиновые волокна (тропомиозин А).
Рис. 4 Электронная микрофотография гладкомышечного
волокна (увеличено в 60 000 раз). Видны актиновые нити (показаны стрелками).
Химический состав мышц колеблется в зависимости
от вида и возраста животного, типа и функционального состояния M. и ряда
др. факторов. Осн. вещества, входящие в состав по-перечнополосатых M. человека
и животных, и их содержание (в % к сырой массе) представлены ниже:
|
Вода
|
72-80
|
|
Плотные вещества
В том числе: |
20-28
|
|
Белки
|
16,5-20,9
|
|
Гликоген
|
0,3-3,0
|
|
Фосфатиды
|
0,4-1,0
|
|
Холестерин
|
0,06-0,2
|
|
Креатин + креатинфос-фат
|
0,2-0,55
|
|
Креатинин
|
0,003-0,005
|
|
АТФ
|
0,25-0,4
|
|
Карнозин
|
0,2-0,3
|
|
Карнитин
|
0,02-0,05
|
|
Анзерин
|
0,09-0,15
|
|
Свободные аминокислоты
|
0,1-0,7
|
|
Молочная кислота
|
0,01-0,02
|
|
Зола
|
1,0-1,5
|
В среднем ок. 75% сырой массы M. составляет
Функциональные особенности поперечнополосатых
Каждая веточка двигательного нерва, иннервирующего
В M. млекопитающих и человека установлено
В поперечнополосатых M. установлено также
Для характеристики сократительной функции
Систематическая усиленная работа M. (тренировка)
Функциональные особенности гладких мышц.
Своеобразие сскратительной функции гладких
Существенное значение для организма имеет
И. И. Иванов.
Скелетные мышцы человека, различные по
Все скелетные, или соматические (от греч.
У человека ок. 500 M., связанных со скелетом.
В суставах шаровидной формы происходят
Наиболее мощные M. размещаются на туловище.
На основе анализа пространственного положения
Патология мыщц. Нарушения сократит, функции
Саркоплазматич. белки скелетных и гладких
Лит.: Энгельгардт В. А., Ферментативные
вода. Осн. количество плотных веществ приходится на долю белков. Различают
белки миофибриллярные (сократительные) - миозин, актин и их комплекс
- актомиозин, тропомиозин и ряд т. н. минорных белков (а и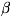 -акти-нины,тропонини
-акти-нины,тропонини
др.), и саркоплазматиче-ские- глобулины X, миогены, дыхательные пигменты,
в частности миоглобин, нуклеопротеиды и ферменты, участвующие в процессах
обмена веществ в M. Из др. соединений важнейшими являются экстрактивные,
принимающие участие в обмене веществ и осуществлении сократительной функции
M.: АТФ, фос-фокреатин, карнозин, анзерин и др.; фосфолипиды, играющие
важную роль в образовании клеточных микроструктур и в обменных процессах;
безазотистые вещества: гликоген и продукты его распада (глюкоза, молочная
к-та и др.), нейтральные жиры, холестерин и др.; минеральные вещества -
соли К, Na
белков - актомиозина, макроэргических соединений, дипепти-дов и др.).
мышц. Поперечнополосатые M. богато снабжены разл. нервами, с помощью к-рых
осуществляется регуляция мышечной деятельности со стороны нервных центров.
Важнейшие из них: двигательные нервы, проводящие к M. импульсы, вызывающие
её возбуждение и сокращение; чувствительные нервы, по к-рым от M. к нервным
центрам поступает информация о её состоянии, и, наконец, адаптационно-трофические
волокна симпатической нервной системы, воздействующие на обмен веществ
и замедляющие развитие утомления M. (см. Адаптационна-трофическая функция).
целую группу мышечных волокон, образующих т. н. моторную единицу, доходит
до отдельного мышечного волокна. Все мышечные волокна, входящие в состав
такой единицы, сокращаются при возбуждении практически одновременно. Под
влиянием нервного импульса в окончаниях двигательного нерва высвобождается
медиатор-ацетилхолин,
взаимодействующий
с холинорецептором пост-синапгической мембраны (см.
Синапсы). В
результате этого происходит повышение проницаемости мембраны для ионов
Na и К, что, в свою очередь, обусловливает её деполяризацию (появление
постсинап-тического потенциала). После этого на соседних участках мембраны
мышечного волокна возникает волна возбуждения (волна электроотрицательности),
к-рая распространяется по скелетному мышечному волокну обычно со скоростью
неск. м в 1 сек. В результате возбуждения M. изменяет свои эластические
свойства. Если точки прикрепления M. не фиксированы неподвижно, происходит
её укорочение (сокращение). При этом M. производит определённую механич.
работу. Если точки прикрепления M. неподвижны, в ней развивается напряжение.
Между возникновением возбуждения и появлением волны сокращения или волны
напряжения протекает нек-рое время, наз. латентным периодом. Сокращение
M. сопровождается выделением тепла, к-рое продолжается в течение определённого
времени и после их расслабления.
существование "медленных" мышечных волокон (к ним принадлежат "красные",
содержащие дыхат. пигмент миоглобин) и "быстрых" ("белых", не имеющих миоглобина),
различающихся скоростью проведения волны сокращения и её продолжительностью.
В "медленных" волокнах млекопитающих длительность волны сокращения примерно
в 5 раз больше, а скорость проведения в 2 раза меньше, чем в "быстрых"
волокнах. Почти все скелетные M. относятся к смешанному типу, т. е. содержат
как "быстрые", так и "медленные" волокна. В зависимости от характера раздражения
возникает либо одиночное - фазное - сокращение мышечных волокон, либо длительное
- тетаническое. Тетанус возникает в случае поступления в M. серии
раздражений с такой частотой, при к-рой каждое последующее раздражение
ещё застаёт M. в состоянии сокращения, вследствие чего происходит суммирование
сократит, волн. H. E. Введенский установил, что увеличение частоты
раздражений вызывает возрастание тетануса, но лишь до известного предела,
наз. им "оптимумом". Дальнейшее учащение раздражений уменьшает тетаническое
сокращение (пессимум). Развитие тетануса имеет большое значение
при сокращении "медленных" мышечных волокон. В M. с преобладанием "быстрых"
волокон макс, сокращение - обычно результат суммации сокращений всех моторных
единиц, в к-рые нервные импульсы поступают, как правило, неодновременно,
асинхронно.
существование т. н. чисто тонических волокон, к-рые особенно широко представлены
в M. земноводных и пресмыкающихся. Тонические волокна участвуют в поддержании
"неутомляемого" мышечного тонуса. Тоническим сокращением наз. медленно
развивающееся слитное сокращение, способное длительно поддерживаться без
значит, энергетич. затрат и выражающееся в "неутомляемом" противодействии
внешним силам, стремящимся растянуть мышечный орган. Тонические волокна
реагируют на нервный импульс волной сокращения лишь локально (в месте раздражения).
Тем не менее, благодаря большому числу концевых двигательные бляшек
тоническое
волокно может возбуждаться и сокращаться всё целиком. Сокращение таких
волокон развивается настолько медленно, что уже при весьма малых частотах
раздражения отдельные волны сокращения накладываются друг на друга и сливаются
в длительно поддерживающееся укорочение. Длительное противодействие тонических
волокон, а также медленных фазных волокон растягивающим усилиям обеспечивается
не только упругим напряжением, но и возрастанием вязкости мышечных белков.
M. пользуются понятием "абсолютной силы", к-рая является величиной, пропорциональной
сечению M., направленной перпендикулярно её волокнам, и выражается в кг/см2.
Так,
напр., абс. сила двуглавой M. человека равна 11,4, икроножной - 5,9 кг/см2.
увеличивает их массу, силу и работоспособность. Однако чрезмерная работа
приводит к развитию утомления, т. е. к падению работоспособности M. Бездеятельность
M. ведёт к их атрофии.
Гладкие M. внутренних органов по характеру иннервации, возбуждения и сокращения
существенно отличаются от скелетных M. Волны возбуждения и сокращения протекают
в гладких M. в очень замедленном темпе. Развитие состояния "неутомляемого"
тонуса гладких M. связано, как и в тонических скелетных волокнах, с замедленностью
сократительных волн, сливающихся друг с другом даже при редких ритмических
раздражениях. Для гладких M. характерна также способность к автоматизму,
т.
е. к деятельности, не связанной с поступлением в 'M. нервных импульсов
из центральной нервной системы. Установлено, что способностью к ритмическому
самопроизвольному возбуждению и сокращению обладают не только нервные клетки,
имеющиеся в гладкий M., но и сами гладко-мышечные клетки
M. позвоночных животных определяется не только особенностями их иннервации
и гистологич. строения, но и спецификой их химич. состава: более низким
содержанием контрактильных белков (актомиозина), макроэргических соединений,
в частности АТФ, низкой АТФ-азной активностью миозина, наличием в них водорастворимой
модификации актомиозина - тоноактомиозина и т. д.
способность гладких мышц изменять длину без повышения напряжения (наполнение
полых органов, например мочевого пузыря, желудка и др.).
форме, величине, положению, составляют св. 40% массы его тела. При сокращении
происходит укорочение M., к-рое может достигать 60% их длины; чем длиннее
M. (самая длинная M. тела - портняжная - достигает 50 см), тем больше
размах движений. Сокращение куполообразной M. (напр., диафрагмы) обусловливает
её уплощение, сокращение кольцеобразных M. (сфинктеров) сопровождается
сужением или закрытием отверстия. M. радиального направления, наоборот,
вызывают при сокращении расширение отверстий. Если M. расположены между
костными выступами и кожей, их сокращение обусловливает изменение кожного
рельефа.
soma - тело), M. по топо-графо-анатомич. принципу могут быть разделены
на M. головы, среди к-рых различают мимические и жевательные M., воздействующие
на нижнюю челюсть, M. шеи, туловища и конечностей. M. туловища покрывают
грудную клетку, составляют стенки брюшной полости, вследствие чего их делят
на M. груди, живота и спины. Расчленённость скелета конечностей служит
основанием для выделения соответствующих групп M.: для верхней конечности
- это M. плечевого пояса, плеча, предплечья и кисти; для нижней конечности
- M. тазового пояса, бедра, голени, стопы.
Среди них одни крупные (напр., четырёхглавая M. бедра), другие - мелкие
(напр., короткие мышцы спины). Совместная работа M. выполняется по принципу
синергизма, хотя отд. функциональные группы M. при выполнении определ.
движений работают как антагонисты. Так, спереди на плече находятся двуглавая
и плечевая M., выполняющие сгибание предплечья в локтевом суставе, а сзади
располагается трёхглавая M. плеча, сокращение к-рой вызывает противоположное
движение - разгибание предплечья.
простые и сложные движения. Напр., в тазобедренном суставе сгибание бедра
вызывает пояснично-подвздошная M., разгибание - большая ягодичная. Бедро
отводится при сокращении средней и малой ягодичных M., а приводится с помощью
пяти M. медиальной группы бедра. По окружности тазобедренного сустава локализуются
также M., к-рые обусловливают вращение бедра внутрь и кнаружи.
Это M. спины - выпрямитель туловища, M. живота, составляющие у человека
особую формацию - брюшной пресс. В связи с вертикальным положением
тела M. нижней конечности человека стали более сильными, поскольку, кроме
участия в локомоции, они обеспечивают опору тела. M. верхней конечности
в процессе эволюции, напротив, сделались более ловкими, гарантирующими
выполнение быстрых и точных движений.
и функциональной деятельности M. совр. наука пользуется также следующим
их объединением: группа M., осуществляющая движения туловища, головы и
шеи; группа M., осуществляющая движения плечевого пояса и свободной верх,
конечности; M. нижней конечности. В пределах этих групп выделяются более
мелкие ансамбли. В. В. Куприянов.
M. и их способности к развитию и поддержанию тонуса наблюдаются при гипертонии,
инфаркте миокарда, мио-дистрофии, атонии матки, кишечника, мочевого пузыря,
при разл. формах параличей (напр., после перенесённого полиомиелита) и
др. Патология, изменения функций мышечных органов могут возникать в связи
с нарушениями нервной или гуморальной регуляции, повреждениями отд. M.
или их участков (напр., при инфаркте миокарда) и, наконец, на клеточном
и субклеточном уровнях. При этом может иметь место нарушение обмена веществ
(прежде всего ферментной системы регенерации макроэргических соединений
- гл. обр. АТФ) или изменение белкового сократит, субстрата. Указанные
изменения могут быть обусловлены недостаточным образованием мышечных белков
на почве нарушения синтеза соответствующих информационных, или матричных,
РНК, т. е. врождённых дефектов в структуре ДНК хромосомного аппарата клеток.
Последняя группа заболеваний, т. о., относится к числу наследственных
заболеваний.
M. представляют интерес не только с точки зрения возможного участия их
в развитии вязкого последействия. Многие из них обладают ферментативной
активностью и участвуют в клеточном метаболизме. При повреждении мышечных
органов, напр, при инфаркте миокарда или нарушении проницаемости поверхностных
мембран мышечных волокон, ферменты (креатинкиназа, лактат-дегидрогеназа,
альдолаза, аминотрансфе-разы и др.) могут выходить в кровь. T. о., определение
активности этих ферментов в плазме крови при ряде заболеваний (инфаркт
миокарда, миопатии и др.) представляет серьёзный клинич. интерес.
и механические свойства белков мышц, -"Успехи современной биологии", 1941,
т. 14, в. 2; С е н т - Д ж и о р д ь и А., О мышечной деятельности, пер.
с англ., M., 1947; Иванов И. И., Юрьев В. А., Биохимия и патобиохимия мышц,
Л., 1961; Поглазов Б. Ф., Структура и функции сократительных белков, M.,
1965; X а и а-ш и Т., Как клетки движутся, в кн.: Живая клетка, пер. с
англ., 2 изд., M., 1966; Хаксли Г., Механизм мышечного сокращения, в сб.:
Молекулы и клетки, пер. с англ., в. 2, M., 1967; Смит Д., Летательные мышцы
насекомых, там же; Бендолл Дж., Мышцы, молекулы и движение, пер. с англ.,
M., 1970; Арронет H. И., Мышечные и клеточные сократительные (двигательные)
модели, Л., 1971; Л ё в и А., С и к е виц Ф., Структура и функции клетки,
пер. с англ., M., 1971; Иванов И- И., Некоторые актуальные проблемы эволюционной
биохимии мышц, "Журнал эволюционной биохимии и физиологии", 1972, т. 8,
№ 3; Gibbons I. R., The biochemistry of motility, "Annual Review of Biochemistry",
1968, v. 37, p. 521. И. И. Иванов.
А Б В Г Д Е Ё Ж З И Й К Л М Н О П Р С Т У Ф Х Ц Ч Ш Щ Ъ Ы Ь Э Ю Я